炒股就看金麒麟分析师研报,权威,专业,及时,全面,助您挖掘潜力主题机会!
我们体内约有36万亿个细胞,从脑细胞到肝细胞、从血细胞到皮肤细胞,每一个都在各司其职、井井有条地工作。[1] 一个神经元从不会分泌胰岛素,一个肝细胞也不会突然产生心跳节律。它们没有大脑、不会思考、没有意志,却能精准地执行其“职责”。
这种精密分工的源头可以追溯到发育的起点——从一颗受精卵开始,那些基因完全相同的细胞,竟能在短短几天内自动分化成器官组织、血液骨骼、神经网络......而这种清晰的分工背后却没有一个中央指挥系统,仿佛与生俱来。
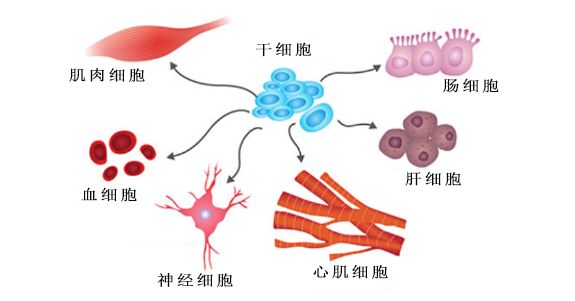 细胞分化示意图
细胞分化示意图(图片来源:Journal of the Royal Society Interface)
所以,细胞没有意识,为何能精准执行生命功能?这不是哲学问题,而是生物学问题。它背后牵涉的是:细胞的命运如何被决定?细胞又是如何“知道”该做什么的?答案隐藏在现代发育生物学和分子生物学的核心原理之中。
我们身体的细胞几乎都含有完全相同的DNA序列,这套“生命说明书”在受精的那一刻就已写定。可为什么它们最终却走上了不同的命运道路?
原因在于,并非所有“生命说明书”里的内容都会在每一个细胞里被读出来使用,而决定哪部分基因被表达的,正是基因表达的调控系统。也就是说,多细胞生命的奥秘,不是不同细胞有不同的基因,而在于它们如何使用相同的基因组,通过选择性表达产生不同的细胞类型。
对于真核细胞构成的个体(例如人类),其基因表达调控至少包含四个部分,层层递进,精妙无比:
染色质结构与表观遗传修饰:DNA并不是裸露存在的,而是缠绕在组蛋白上形成染色质。紧密缠绕的区域(异染色质)通常关闭表达,而松散区域(常染色质)更容易被转录。表观遗传修饰是指是指在不改变DNA序列的前提下,通过可逆的化学修饰(如DNA甲基化、组蛋白修饰或RNA修饰)调控基因表达的一种遗传调控机制。其中,组蛋白乙酰化会让染色质结构变松,从而促进基因激活,而DNA的甲基化则往往意味着沉默表达。[2][3] 这就像同一本书,有些章节被封存,有些则可以随时翻开,决定了基因可读性。
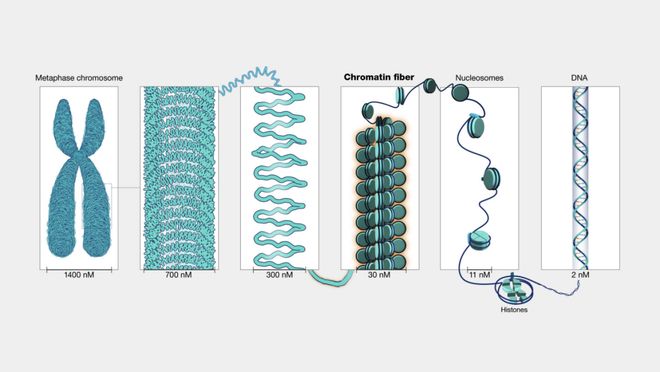
染色体结构尺度与组成
(图片来源:NHGRI)
顺式调控元件与转录因子:每个基因前都有启动子作为基本开关,而增强子与沉默子则像是远程遥控器,负责增强或压制表达强度。转录因子就是负责读取这些模块并下达命令的蛋白。
RNA水平的调控:即便一个基因被转录生成了mRNA,这条信息也可以被剪接成不同的版本,产生不同的蛋白质。此外,miRNA等非编码RNA的参与,也能严密调控mRNA的稳定性、翻译效率。
蛋白翻译后的修饰与命运:蛋白质在核糖体上完成翻译后,并不会立刻具备完整的生物功能,而是需要经历一系列被称为翻译后修饰的过程来决定其最终“命运”。这些修饰包括如泛素化,即在特定位点连接泛素分子,通常标志该蛋白将被细胞内的蛋白酶体系统识别并降解;磷酸化则是通过激酶将磷酸基团加到丝氨酸、苏氨酸或酪氨酸残基上,从而改变蛋白的构象、电荷状态或与其他分子的结合能力,使其激活、抑制或重新定位;此外,许多蛋白还需通过构象折叠、伴侣蛋白辅助、剪切加工等方式获得成熟结构,才能行使特定功能。正是这些翻译后修饰机制,使细胞能够在不改变基因编码的前提下,灵活调控蛋白功能、寿命和信号反应,是维持生理稳态与应答环境变化的重要调控手段。[4]
由此观之,基因表达并非随心所欲,而是像一道多级审批流程,需要层层审核、批准、调控。
虽然细胞拥有相同的DNA,但通过层层调控系统,它们只能打开属于自己那部分章节。那么,是哪一刻、在什么条件下,一个细胞会不可逆地走向特定分化命运?这正是细胞命运决定机制的核心问题。
在胚胎发育早期,所有细胞都是全能干细胞,也就是什么都能做的小胚胎。但这种全能性仅存在于受精后4—8细胞期(胚胎发育的一个特定阶段,指受精卵经过多次分裂后形成4个至8个细胞的时期)。进入囊胚期后,这些细胞就变为多能干细胞,开始进入命运的分岔路口,逐步锁定自己的“职业路径”。
这个命运分配过程,受到一系列高度保守且精妙的调控机制控制:
形态发生因子:这些是胚胎发育过程中形成浓度梯度的关键信号分子。常见的有Shh、Wnt、BMP等多种信号,细胞通过感知这些信号的局部浓度差异,找到自己所处的位置,从而激活不同的命运程序。例如,Shh高浓度诱导神经管腹侧发育出运动神经元;[5]Wnt的前后梯度决定胚体头尾轴的分化;[6]BMP活跃区域趋向表皮,而其抑制区域则诱导神经组织的生成。
主调转录因子:细胞在接受信号刺激后,会启动某些关键转录因子,它们能一锤定音地定义细胞身份。比如MyoD决定肌肉细胞[7]、Pax6主导眼睛发育[8],而PU.1和GATA1之间则决定免疫系统中不同细胞类型的分化[9]。这些关键因子就像是为每一个细胞分配着未来的角色。
为了确保细胞命运决定的不可逆性,大多数分化网络中还存在互斥机制与正反馈系统。一旦某个路径被激活,它就会增强自身表达,并压制其他可能性,最终锁定命运方向。
一旦细胞的命运被锁定,它便需要一整套内部操作系统来长期执行——这就是基因调控网络(Gene Regulatory Network, GRN)。GRN不是单一开关,而是一个由转录因子、调控元件、靶基因等组成的复杂、动态运行的信息处理系统。不同的节点彼此激活、互相抑制,构成正负反馈,能够对外部信号快速应答,又在内部维持稳态[10]。
例如,在肝细胞中,当血糖水平升高时,GRN会激活糖原合成基因来调节血糖,这正是其动态响应的体现。与此同时,肝细胞还依赖HNF1α、HNF4α、FoxA、C/EBPβ等多个肝特异性因子的互相激活,排斥非肝谱系基因,从而长期维持其肝细胞的特性。
前文提到,细胞命运具有不可逆性。那是否意味着其命运一旦确定,就不可被更改?实则不然。2006年,日本科学家山中伸弥用四个转录因子,成功地将成年小鼠的皮肤细胞逆转为类似胚胎干细胞的状态,这一突破性成果被称为诱导多能干细胞(iPSC)。这项研究发表于《Cell》杂志[11],并促成他于2012年获得诺贝尔奖。
iPSC的意义在于它首次向世界证明了:细胞命运走向并非单行道,基因调控网络也并非不可破解。只要找到合适的控制点,就可以改写细胞的命运——皮肤细胞可以变为干细胞,再重新分化成神经元、心肌细胞或胰岛β细胞。这表明我们第一次拥有了改变自身细胞命运的强大能力。
拥有了这些能力之后,我们可以做到很多之前无法想象的事情,比如治疗用iPSC技术和细胞重编程方法,我们开始尝试治疗之前被认为需要终身用药的糖尿病。
比如就在今年(2025年),殷浩教授团队联合中国科学院分子细胞科学卓越创新中心程新教授团队,在国际学术期刊《细胞发现》(Cell Discovery)在线发表了最新研究成果。这是国际上首次利用干细胞来源的自体再生胰岛移植,成功治愈胰岛功能严重受损糖尿病的病例报道。[12]
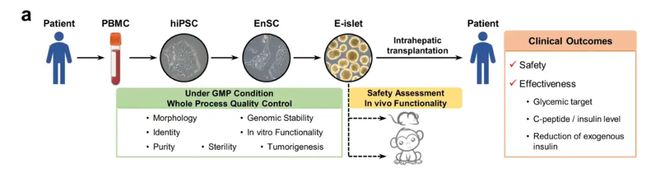
这个治疗流程就是先抽取患者的外周血,分离出外周血单个核细胞(PBMC),作为细胞重编程的起始材料;随后通过引入特定转录因子,将PBMC诱导为具有多向分化潜能的人诱导多能干细胞(hiPSC);在特定诱导条件下,hiPSC被定向分化为人类内胚层干细胞(EnSC),作为构建胰腺细胞系的重要中间前体;经胰腺谱系定向诱导后,EnSC进一步分化成熟为具备葡萄糖响应性胰岛素分泌功能的工程化胰岛细胞团,也就是E-胰岛(engineered islets, E-islet),可作为功能性胰岛替代物用于后续的体内移植与胰岛功能重建。最后把这些具有功能性的E-胰岛植入到患者的肝脏内,就使得患者再次拥有了自身份泌胰岛素的功能。
这不得不说是再生医学领域的一个重要的突破,为一些糖尿病患者提供了新的治愈希望。
看到这里,最初的问题有了回答——单个细胞的确没有意识,其命运由遗传程序与环境信号协同调控决定。但新的问题也随之产生,这些细胞无意识,那我们的意识又是从何而来的?
一个神经元没有思想,它只是按照信号兴奋和传导。但当数百亿神经元通过复杂拓扑连接与递归回路协同活动时,就出现了系统层面的涌现——即意识[13]。认知神经科学中的全球神经元工作空间(GNW)理论提出:只有当信息被“点燃”并在长程回路中广泛广播时,才进入可报告的意识状态[14]。
这就是复杂系统科学中所说的“涌现”:单个水分子不具备湿的属性,但大量水分子聚在一起就能表现出湿的感觉;单只蚂蚁无法完成复杂工作,但蚁群可以协调调配资源、完成惊人的结构建设。同样地,细胞本身没有意识,但它们的集合最终涌现出具有思考能力的我们。
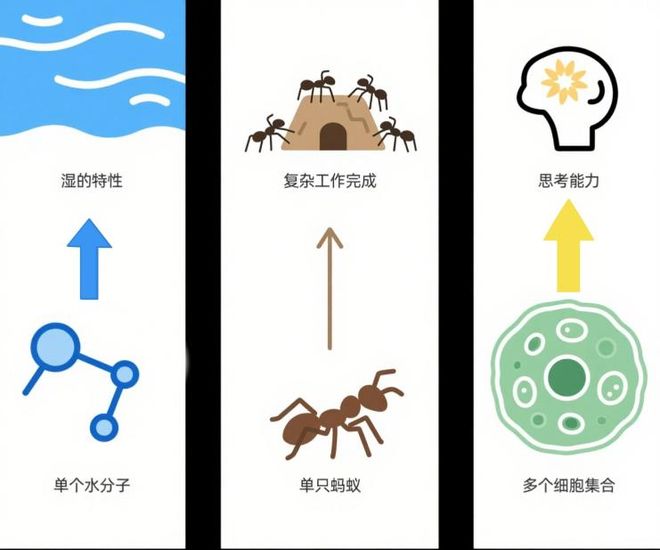
“涌现”的概念
(图片来源:作者使用AI生成)
最近几年,国内外各大生成式大语言模型十分火热,它们也是经历了类似的过程,才能具备一定能力。我们发现,从小模型到大模型,当模型突破某个规模时,它的语言理解能力、生成能力、逻辑推理能力有了极大幅度的非线性提升,这也被认为是AI智慧的“涌现”[15]。
意识作为一种复杂系统的“涌现”现象,其特点在于无法简单地还原为构成部分的属性,而是系统整体协同作用的结果,通常伴随着全新的、不可预测的行为和能力。
从认知神经科学角度看,意识的产生需要大脑以高度复杂的结构进行信息整合。这包括多个脑区间的信息同步、整合记忆重构与未来预测的时序处理回路、反馈与前馈机制之间的联动以及足够高的信号通量来支撑持续的神经活动。这些都是神经网络层级才能具备的功能,远非单一细胞能承载。
所以,细胞之所以没有意识,是因为它没有能产生意识的结构系统;但它们通过各自的程序化行为,最终组成了一个个有意识、会思考的人类个体。细胞无法自我理解,但它们依靠集体运作,赋予了我们认识自我的能力。而对这种协作机制的深入理解,正不断推动着我们在再生医学、疾病治疗乃至人工智能等前沿领域不断突破。
参考文献:
[1] I.A. Hatton, E.D. Galbraith, N.S.C. Merleau, T.P. Miettinen, B.M. Smith, & J.A. Shander, The human cell count and size distribution, Proc. Natl. Acad. Sci. U.S.A. 120 (39) e2303077120, https://doi.org/10.1073/pnas.2303077120 (2023).
[2] Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002 Jan 1;16(1):6-21. doi: 10.1101/gad.947102. PMID: 11782440.
[3] Shogren-Knaak M, Ishii H, Sun JM, Pazin MJ, Davie JR, Peterson CL. Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science. 2006 Feb 10;311(5762):844-7. doi: 10.1126/science.1124000. PMID: 16469925.
[4] Ciechanover, A. Intracellular protein degradation: from a vague idea thru the lysosome and the ubiquitin–proteasome system and onto human diseases and drug targeting.Cell Death Differ12, 1178–1190 (2005). https://doi.org/10.1038/sj.cdd.4401692
[5] Yuichi Nishi, Xiaoxiao Zhang, Jieun Jeong, Kevin A. Peterson, Anastasia Vedenko, Martha L. Bulyk, Winston A. Hide, Andrew P. McMahon; A direct fate exclusion mechanism by Sonic hedgehog-regulated transcriptional repressors. Development 1 October 2015; 142 (19): 3286–3293. doi: https://doi.org/10.1242/dev.124636
[6] Yamaguchi TP. Heads or tails: Wnts and anterior-posterior patterning. Curr Biol. 2001 Sep 4;11(17):R713-24. doi: 10.1016/s0960-9822(01)00417-1. PMID: 11553348.
[7] Davis RL, Weintraub H, Lassar AB. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell. 1987 Dec 24;51(6):987-1000. doi: 10.1016/0092-8674(87)90585-x. PMID: 3690668.
[8] Gehring WJ. The master control gene for morphogenesis and evolution of the eye. Genes Cells. 1996 Jan;1(1):11-5. doi: 10.1046/j.1365-2443.1996.11011.x. PMID: 9078363.
[9] Zhang P, Zhang X, Iwama A, Yu C, Smith KA, Mueller BU, Narravula S, Torbett BE, Orkin SH, Tenen DG. PU.1 inhibits GATA-1 function and erythroid differentiation by blocking GATA-1 DNA binding. Blood. 2000 Oct 15;96(8):2641-8. PMID: 11023493.
[10] E.H. Davidson, & M.S. Levine, Properties of developmental gene regulatory networks, Proc. Natl. Acad. Sci. U.S.A. 105 (51) 20063-20066, https://doi.org/10.1073/pnas.0806007105 (2008).
[11] Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006 Aug 25;126(4):663-76. doi: 10.1016/j.cell.2006.07.024. Epub 2006 Aug 10. PMID: 16904174.
[12] Wu, J., Li, T., Guo, M.et al.Treating a type 2 diabetic patient with impaired pancreatic islet function by personalized endoderm stem cell-derived islet tissue.Cell Discov10, 45 (2024). https://doi.org/10.1038/s41421-024-00662-3
[14] Mashour GA et al. Conscious processing and the GNW hypothesis. Nat Rev Neurosci. 2020;21:751-764.
[15] Wei J et al. Emergent Abilities of Large Language Models. arXiv:2206.07682 (2022).
出品:科普中国
作者:冷哲(科普创作者)
(转自:网易科技)